

Evolucija čovjeka

Evolucija čovjeka podrazumijeva evolutivni proces koji je tijekom vremena doveo do pojave anatomski modernog čovjeka. Čovječja je evolucija u biti dio evolucije primatâ, točnije razvoja roda Homo i pojave vrste Homo sapiens kao jedne od vrsta hominida ili velikih čovjekolikih majmuna. Izučavanje evolucije čovjeka obuhvaća niz različitih znanstvenih disciplina: paleontologiju, antropologiju, arheologiju, jezikoslovlje, embriologiju i genetiku.[1]
Evolucija primatâ počinje u kasnoj kredi. Prema izvršenim genetičkim studijama, primati su se počeli odvajati od ostalih sisavaca prije 85 milijuna godina, a prvi sigurni fosili primatâ pojavljuju se u paleocenu prije 55 milijuna godina.[2] Rod hominida ili velikih čovjekolikih majmuna odvaja se od roda Hylobatidae u razdoblju od 15-20 milijuna godina, a prije 14 milijuna godina Ponginae ili orangutani odvajaju se od roda Hominidae.[3]
Dvonožno je hodanje ili bipedalizam najvažnija prilagodba tribusa hominina. Smatra se da su prvi dvonožni hominini bili Sahelanthropus tchadensis ili Orrorin, dok se potpuno dvonožni Ardipithecus pojavio nešto kasnije. Preci gorila i čimpanza odvajaju se otprilike u istom razdoblju, tako da su Sahelanthropus ili Orrorin vjerojatno naši posljednji zajednički biološki preci. Prvi dvonožni majmuni najzad evoluiraju u rod Australopithecines i poslije u rod Homo.
Prva dokumentirana vrsta roda Homo bio je Homo habilis koji se razvio prije otprilike 2,3 milijuna godina. Homo habilis prva je vrsta kod koje je sa sigurnošću dokazano korištenje kamenog oruđa. Volumen mozga prvih hominina bio je otprilike jednak onom suvremenih čimpanza. Tijekom daljnjih vremenskih razdoblja došlo je do razvoja mozga i povećanja njegova volumena, tzv. procesa encefalizacije. Fosilni ostaci dokazuju da se s pojavom vrste Homo erectus volumen neurokranija udvostručio u odnosu na prve hominine i dosegao 850 cm3.[4] Homo erectus i Homo ergaster bili su prvi hominini koji su napustili Afriku te su se osim Afrike raširili i u Aziju i Europu prije 1,8-1,3 milijuna godina. Pretpostavlja se da su to bile prve vrste koje su koristile vatru i složeno oruđe. Po posljednjoj paleoantropološkoj teoriji recentnog afričkog porijekla (engl. Recent African Ancestry theory) poznatoj i kao teoriji napuštanja Afrike (engl. Out of Africa theory), anatomski su se moderni ljudi razvili u Africi vjerojatno iz vrste Homo heidelbergensis i napustili Afriku prije 100.000 do 50.000 godina, potisnuvši tako lokalne populacije vrsta Denisovskog čovjeka, Homo neanderthalensis (neandertalac), Homo floresiensis te druge.[5][6][7][8][9]
Prvi predstavnik anatomski modernog čovjeka bio je tzv. arhajski Homo sapiens koji se razvio u razdoblju prije 400.000 do 250.000 godina kad je populacija neandertalaca postupno opadala. Suvremeni genetički dokazi ukazuju da više haplotipa u genskom kodu sadašnjih ljudi koji nisu iz Afrike vuče podrijetlo iz DNA neandertalaca i drugih hominina, kao npr. "denisovskog čovjeka". Otprilike 6 % sadašnjeg genskog koda neafričkih naroda mogu se pripisati drugim vrstama hominina, naročito neandertalcima, dokazujući tako da je dolazilo do križanja različitih vrsta roda Homo.[10][11][12]
Anatomski moderni ljudi razvijaju se iz arhajske vrste Homo sapiens u srednjem paleolitiku, otprilike prije 200.000 godina.[13] Prijelaz na modernitet u ponašanju s pojavom uporabe simbola, jezika i specijaliziranog kamenog oruđa dogodio se prije najkasnije 50.000 godina iako postoje hipoteze da se moderno ponašanje možda pojavilo već s pojavom anatomski modernih ljudi.[14]

Evolucija čovjeka okarakterizirana je nizom promjena u anatomskom izgledu, fiziologiji i ponašanju, koji su se postupno ustalili od odvajanja predaka čovjeka od predaka ostalih majmuna. Najvažnije evolutivne promjene su dvonožno hodanje ili bipedalizam, povećanje moždanog volumena ili encefalizacija, produženje trudnoće i smanjenje anatomskih razlika među spolovima. Međusobne interakcije između navedenih promjena još su uvijek predmet rasprava.[15]
Još se jedna bitna prilagodba ustalila tijekom evolucije čovjeka, a to je čvrstoća i preciznost stiska šake, koja se prvi put razvila kod Homo erectusa.[16]
Dvonožno hodanje ili bipedalizam jedna je od temeljnih prilagodbi tribusa hominina i smatra se osnovnim uzrokom niza promjena u anatomskom izgledu kostura čovjeka. Smatra se da su prvi dvonožni hominini bili Sahelanthropus tchadensis[17] i Orrorin, dok je Ardipithecus bio prvi isključivo dvonožni hominin. Majmuni koji hodaju na šakama kao gorile i čimpanze, odvojili su se kao zasebna skupina otprilike u isto doba, tako da su Sahelanthropus ili Orrorin vjerojatno naši posljednji zajednički preci. Prvi dvonožni hominini su se najzad razvili u rod Australopithecines, a kasnije u rod Homo. Postoje brojne hipoteze glede korisnosti dvonožnog hodanja; od oslobađanja gornjih udova za dosezanje i nošenje hrane, čuvanja energije tijekom hodanja, olakšavanja trčanja na duge staze do smanjenja zagrijavanja tijela smanjenjem površine tijela koja je izložena Suncu.
Razvoj dvonožnog hodanja anatomski je promijenio čitav kostur, ne samo noge i zdjelice, već i kralježnicu, stopala, gležnjeve i lubanju. Najvažnije promjene desile su se u području zdjelice, gdje se bočna kost ili ilium bitno skratila i proširila kako bi se baricentar tijela održao stabilnim za vrijeme hodanja. Skraćenje zdjelice razvilo se kao prilagodba dvonožnom hodanju te je imalo važne posljedice na mehanizam ljudskog poroda, koji je kod anatomski modernih ljudi postao teži u odnosu na ostale primate. Bedrena se kost razvila tako da se premjestila pod kut u odnosu na zdjelicu, kako bi se baricentar tijela približio geometrijskom centru tijela. Koljena i kukovi su ojačali kako bi podnijeli težinu trupa, a kralježnica je poprimila oblik dvostrukog slova S, dobivši tako učinak amortizera pri hodanju i trčanju. Slabinski su kralješci postali kraći i širi. Došlo je i do promjena na stopalu; nožni se palac izravnao s ostalim prstima kako ne bi smetao pri hodanju. Gornji su se udovi skratili u odnosu na donje olakšavši na taj način trčanje. Uspravnim položajem trupa, ''foramen magnum'' se pomakao na donju stranu lubanje.
Kod Homo sapiensa, ali i kod drugih vrsta roda Homo, razvio se mozak puno većeg volumena nego kod ostalih primata. Prosječni volumen mozga kod Homo sapiensa je 1330 cm3, što je više nego dvostruki volumen u odnosu na mozak čimpanza i gorila.[18] Razvoj mozga ili encefalizacija započeo je s Homo habilisom, koji je sa svojih 600 cm3moždanog volumena bio sličan današnjim čimpanzama. Najveći volumen mozga, oko 1200–1900 cm3, dosegao je neandertalac, što je više nego kod suvremenog čovjeka. Postoje bitne razlike u strukturi i načinu razvoja ljudskog mozga u odnosu na ostale majmune, ne samo u dimenzijama. Poseban slijed razvoja ljudskog mozga omogućavaju duga razdoblja učenja tijekom djetinjstva i učenje jezika.[19][20][21][22]
Povećanje moždanog volumena obuhvatilo je neke dijelove mozga u većoj mjeri nego druge. Slijepoočni režanj (lobus temporalis) se izuzetno puno razvio, budući se u njemu nalaze centri za kontrolu govora, a isto se tako razvio i prefrontalni režanj koji kontrolira složene procese odlučivanja i društveno ponašanje.[18] Encefalizacija je vezana za povećanu količinu mesa u prehrani,[23][24] i za pojavu kuhanja namirnica.[25] Pretpostavlja se da je pojačani razvoj mozga bio potaknut i time što su zajednice ljudi postale vrlo složene, te je razvoj inteligencije postao važan kao instrument rješavanja problema unutar zajednice.
Kod čovjeka, anatomske razlike među spolovima uglavnom su umanjene u odnosu na ostale primate. To je vidljivo u smanjenoj veličini očnjaka kod muškarca čovjeka u odnosu na mužjake drugih primata, kao i u smanjenom čeonom grebenu kod čovjeka i općem smanjenju fizičke konstitucije. Još jedna uočljiva razlika u fiziologiji ljudske spolnosti naspram ostalih primata je razvoj tzv. skrivene ovulacije. Čovjek je jedini pripadnik primata u kojem su žene plodne tijekom cijele godine i kod kojih ne dolazi do vidljivih znakova na tijelu za vrijeme plodnih dana kao npr. nabreknuća genitalija. Pored toga, kod čovjeka postoje razlike u sekundarnim spolnim odlikama, poput različitog rasporeda dlaka i potkožne masti te fizičke konstitucije koja je oko 25% krupnija kod muškaraca u odnosu na žene. Smanjena razlika među spolovima interpretira se kao prilagodba potrebi jačeg vezanja parova i suradnji pri podizanju potomstva s dugim djetinjstvom.

Riječ homo, koja je naziv za biološki rod kojem pripadaju ljudi dolazi iz iste latinske riječi što znači "čovjek". Prvi ju je koristio poznati švedski taksonom Carl Linné u svom sustavu klasifikacije živih bića. Linné i ostali taksonomi onog doba s pravom su smatrali da su veliki čovjekoliki majmuni najbliži srodnici čovjeka zbog morfoloških i anatomskih sličnosti.
Mogućnost da su ljudi i majmuni imali zajedničke pretke postaje jasnija tek nakon 1859. s objavom knjige "Podrijetlo vrsta" engleskog biologa Charlesa Darwina. Ovo je znanstveno djelo argumentiralo evoluciju novih vrsta od starijih putem prirodnog odabira, ali se nije doticalo pitanje evolucije čovjeka. Jedini spomen o tom pitanju je rečenica: "Bacit će se svijetlo na podrijetlo čovjeka i njegovu povijest."
Prve zabilježene rasprave o podrijetlu čovjeka bile su one između Thomasa Huxleya i Richarda Owena. Huxley je zastupao tezu evolucije čovjeka od majmuna argumentirajući je mnogim morfološkim sličnostima što je i obrazložio 1863. u svojoj knjizi "Dokaz čovjekovog mjesta u prirodi" (eng.: Evidence as to Man's Place in Nature). Međutim, mnogi Darwinovi istomišljenici onog doba nisu se slagali s idejom da se čovjekove umne mogućnosti i moralna osjetljivost mogu objasniti prirodnim odabirom. Darwin je primijenio teoriju evolucije i spolne selekcije na čovjeka u svojoj drugoj knjizi "Podrijetlo čovjeka" (eng.: The Descent of Man, and Selection in Relation to Sex) objavljenoj 1871.[26]

Najveći problem onoga doba bio je manjak fosilnih ostataka ljudskih predaka. Prvo bitno otkriće fosila hominida desilo se 1891. kad je francuski arheolog Eugène Dubois otkrio ostatke "Javanskog pračovjeka" blizu mjesta Trinil na otoku Javi. Danas je taj pračovjek klasificiran kao Homo erectus. Tek je dvadesetih godina prošlog stoljeća otkrivena veća količina različitih fosilnih ostataka u Africi, gradeći postupno sliku ljudskih međuvrsta. Raymond Dart je 1925. opisao prvog Australopitecusa africanusa, "dijete iz Taunga". Pronađeni fosilni ostaci otkriveni su u pećini te su se sastojali od dijela lubanje, donje čeljusti i fosiliziranog odljeva mozga (vidi sliku). Iako mu je moždani volumen bio relativno malen (410 cm3), oblik mozga je bio zaobljen što je karakteristično za modernog čovjeka. Kod pronađenih ostataka vidljivi su kratki očnjaci i foramen magnum pozicioniran s donje strane lubanje, što je znak uspravnog držanja i dvonožnog hodanja. Navedeni dokazi uvjerili su Darta da je dijete iz Taunga dvonožni ljudski predak, tranzicijski oblik između ljudi i majmuna.
Tijekom šezdesetih i sedamdesetih godina prošlog stoljeća pronađene su stotine fosilnih ostataka hominida, pogotovo u istočnoj Africi. Najvažnija fosilna nalazišta onog doba bila su klanac Olduvai i jezero Turkana. Najpoznatiji i najplodniji istraživači i paleoantropolozi onog doba bila je obitelj Leakey: Louis Leakey, njegova žena Mary Leakey a kasnije i njihov sin Richard i snaha Meave. Iz fosilnih nalazišta Olduvaija i Turkane prikupili su mnoštvo fosila roda Asutralopithecines, prve pripadnike roda Homo i ostatke Homo erectusa. Ova otkrića potvrdila su Afriku kao kolijevku ljudskog roda. U osamdesetim godinama prošlog stoljeća Etiopija je postala najvažnije paleoantropološko nalazište nakon što je Don Johanson pronašao "Lucy", najbolje očuvanog fosila vrste Australopithecus afarensis u blizini mjesta Hadar u sjevernoj Etiopiji.
U tom će se području kasnije otkriti mnoštvo fosila različitih hominina, pogotovo od strane paleontologa Tim Whitea koji će u devedesetim godinama pronaći fosile Ardipithecusa ramidusa.
Genetska "revolucija" u proučavanju evolucije čovjeka započela je kad su biolozi Vincent Sarich i Allan Wilson izmjerili intenzitet imunosnih reakcija na albumin u krvnom serumu između različitih vrsta životinja, uključujući ljude i čovjekolike majmune poput čimpanzi i gorila. Snaga imunosne reakcije može se brojčano izraziti kao "imunološka udaljenost" (eng.: "Immunological Distance"), koja je razmjerna razlikama u sekvenciji aminokiselina između homolognih bjelančevina kod različitih vrsta.
Nakon što su odredili kalibracijsku krivulju između parova vrsta i poznatog vremena divergencije fosilnih nalaza, podaci su se mogli koristiti kao molekularni sat za procjenu vremena razdvajanja parova kod siromašnijih ili nepoznatih fosilnih nalaza. U članku objavljenom 1967. u časopisu "Science" Sarlich i Wilson su procijenili vrijeme razdvajanja ljudi i majmuna na četiri do pet milijuna godina. U to se vrijeme pretpostavljalo da je vrijeme razdvajanja ljudi i majmuna desilo pred 10 do 30 milijuna godina. Kasniji fosilni nalazi, pogotovo otkriće "Lucy", i pomnije datiranje starijeg fosilnog materijala (Ramapitecusa), dokazali su da se vrijeme divergencije zbilo kasnije, potvrdivši tako valjanost imunološke metode. Primjena imunološke metode revolucionirala je istraživanja u molekularnoj evoluciji.
Tijekom devedesetih godina prošlog stoljeća više skupina paleoantropologa vršilo je istraživanja duž cijele Afrike u potrazi za najstarijim foslinim dokazima razdvajanja hominina od ostalih čovjekolikih majmuna. Meave Leakey je 1994. otkrila prve fosile Australopithecusa anamensisa, ali grupa Tima Whitea svojim otkrićem Ardipithecusa ramidusa pomiče dokaze divergencije ljudi i majmuna na 4,2 milijuna godina. Martin Pickford i Brigitte Senut 2000. otkrivaju 6 milijuna godina stare fosile dvonožnog hominina na brdima Tugen u Keniji kojega su nazvali Orrorin tugenensis. Grupa paleoantropologa predvođena Michelom Brunetom 2001. pronalaze fosilnu lubanju Sahelanthropusa tchadensisa čija je starost procijenjena na 7,2 milijuna godina. Brunet pretpostavlja da je Sahelanthropus bio dvonožan, te da stoga spada u tribus hominina.

Antropolozi su 1980-ih godina bili duboko podijeljeni oko činjenica u vezi migracija i reproduktivnih barijera roda Homo. Znanstvenici su se kasnije poslužili napretkom u genetici kako bi istražili i riješili ova otvorena pitanja. Prema posljednjim dokazima, pripadnici roda Homo emigrirali su izvan Afrike bar tri puta. Najprije je to učinio Homo erectus, pa Homo heidelbergensis i na posljetku Homo sapiens, s naznakama i nekih novijih migracija (npr. afro-azijska jezična porodica na Bliskom Istoku).
Prema evolucijskom modelu nedavnog afričkog podrijetla modernog čovjeka (teorija „Out of Africa“), moderni se Homo sapiens definirao kao vrsta u Africi prije oko 200 000 godina i njegove kasnije migracije duž cijele Euroazije rezultirale su nestankom ostalih vrsta roda Homo. Ovaj su evolucijski model razvili Chris Stringer i Peter Andrews.[28][29]
Konkurentska hipoteza multiregionalne evolucije pretpostavljala je da se cijeli rod Homo sastojao od jedne velike međusobno povezane populacije (kao što je to danas slučaj), koja je simultano i konstantno evoluirala na više mjesta u svijetu u rasponu od par milijuna godina. Ova je hipoteza stoga odbijala postojanje različitih vrsta roda Homo. Model multiregionalne evolucije je prvi predložio Milford H. Wolpoff 1988. godine.[30][31]
Postignuti napredak u tehnici skvenciranja DNK-a, pogotovo u mitohondrijskog DNK-a i kasnije u DNK-u kromosoma Y uvelike je unaprijedio spoznaje o podrijetlu čovjeka.[32][33][34]
Sekvencirani su uzorci mtDNK i Y-DNK kod domorodačkog stanovništva iz cijelog svijeta, koji su otkrili informacije o genetskom naslijeđu muške i ženske populacije.[35] Rezultati dobiveni usporedbom razlika u genetskom stablu analiziranih uzoraka interpretirani su kao čvrsti dokaz teorije nedavnog pojedinačnog podrijetla (teorije „Out of Africa“).[36] Uzorci su pokazali najveću raziličitost upravo kod afričkog domorodačkog stanovništva, potvrđujući tako ideju da je Afrika genetska kolijevka prvih žena i muškaraca vrste Homo sapiens.[37] Proučavanja mitohondrijskog DNK-a i DNK-a kromosoma Y dalo je čvrste dokaze u korist teoriji nedavnog afričkog pretka. Analizirajući genealoška stabla sastavljena od 133 vrste mtDNK-a, stručnjaci su dokazali da je ljudski rod genetski potekao od jednog ženskog afričkog pretka, kojeg su nazvali mitohondrijskom Evom.[38]
Široka studija afričkih genetskih različitosti sprovedena pod vodstvom Sarah Tishkoff, dokazala je da narod San ima najveću genetsku različitost između 113 testiranih različitih populacija, što ga čini jednim od 14 drevnih populacijskih klastera. Istraživanje je odredilo mjesto početka migracija modernih ljudi u jugozapadnu Afriku, u blizini obalne granice država Namibije i Angole.[39]Richard Leakey je imao premalo fosilnih dokaza da bi riješio ovo pitanje.[40] Studije haplogroupa na DNK kromosoma Y mitohondrijskog DNK-a čvrsto su podržale teoriju nedavnog afričkog podrijetla.[41] Dokazi iz analiza autosomske DNK također uvelike podržavaju istu teoriju.
Neke studije, međutim, sugeriraju da postoje dokazi miješanja arhajskog homo sapiensa s modernim ljudima.[42]
Nedavno sekvenciranje genoma neandertalca[43] i denisovskog čovjeka[44] dokazuje da je došlo do određenog stupnja genetskog miješanja. Moderni ljudi izvan Afrike imaju 2 – 4 % alela neandertalaca u svom genomu, a neki narodi Melanezije imaju dodatnih 4 – 6 % alela denisovskog čovjeka.
Ovi novi rezultati nisu u kontradikciji s modelom nedavnog afričkog pretka, osim u svojoj najstriktnijoj interpretaciji. Nakon oporavka od genetskog uskog grla koji je možda bio prouzročen katastrofom izazvanom eksplozijom supervulkana Toba, relativno mala grupa ljudi napušta Afriku i nakratko se genetski križa s neandertalcima, vjerojatno na području Bliskog istoka ili možda već u sjevernoj Africi prije daljnje migracije. Njihovi još uvijek dominantno afrički potomci šire se u druge krajeve i naseljavaju cijeli svijet. Jedan dio ovih ljudi dolazi u kontakt s denisovskim ljudima i miješa se s njima, vjerojatno na području jugoistočne Azije, te kasnije nastanjuje Melaneziju.[45] HLA haplotip podrijetlom od neandertalca i denisovskog čovjeka pronađen je u modernim populacijama Euroazije i Oceanije.[12][46]
Postoje još uvijek divrgentne hipoteze da li je došlo do samo jedne ili do više različitih migracija. Teorijski model višestrukog širenja uključuje i teoriju južnog širenja,[47] koju zadnjih godina sve više podržavaju genetski, lingvistički i arheološki dokazi. Po toj teoriji došlo je do obalne migracije modernih ljudi iz područja Roga Afrike prije oko 70 000 godina. Ova je grupa pridonijela naseljavanju jugoistočne Azije i Oceanije, objašnjavajući tako otkriće ranih ljudskih naseobina u tim područjima puno prije nego što na Levantu. Drugi val migracija modernih ljudi proširio se kroz poluotok Sinaj u Aziju, što je imalo kao rezultat formiranje većine ljudske populacije Euroazije. Ova druga grupa ljudi razvila je sofisticiranu tehnologiju izrade kamenog alata i bila je manje ovisna o izvorima hrane koja se nalazi uz morsku obalu od prve grupe migranata. Velik dio dokaza migracija prve grupe ljudi vjerojatno je uništen podizanjem razine mora na kraju svakog glacijalnog maksimuma.[47] Model višestrukih migracija je u kontradikciji sa studijama koje dokazuju da sve populacije jugoistočne Azije i populacije Oceanije potječu iz iste linije mitohondrijskog DNK-a, što podržava teoriju jednog jedinog migracijskog vala iz Afrike od kojeg potječu sve ostale neafričke populacije.[48]
Znanstveni dokazi ljudske evolucije proizlaze iz nekoliko znanstvenih disciplina. Osnovni izvor spoznaje o ljudskom evolutivnom razvoju je tradicionalno bilo sakupljanje fosilnih dokaza, ali s razvojem genetike od 1970-ih na dalje analiza DNK-a postala je jednako važan izvor informacija. Izučavanje ontogeneze, filogeneze i evolucijske biologije i kod kralježnjaka i kod beskralježnjaka daje nam važan uvid u evoluciju života na Zemlji. Znanstvena disciplina koja se bavi evolutivnim razvojem čovjeka naziva se paleoantropologija.[49]

Dvije životinjske vrste koje su evolutivno najbliže čovjeku su gorile i čimpanze.[50]
Sa završetkom sekvenciranja ljudskog genoma i genoma čimpanze, sadašnja je procjena da se sličnost ovih dvaju genoma kreće od 95% do 99%.[50][51][52] Koristeći tehniku tzv. molekularnog sata koji procjenjuje vrijeme potrebno za akumuliranje određenog broja divergentnih mutacija između dvije vrste, može se izračunati približni vremenski period u prošlosti u kojem je došlo do razdvajanja dviju vrsta. Giboni (porodica Hylobatidae) i orangutani ( rod Pongo) bili su prve grupe koje su se odvojile od evolucijske linije koje vode do čovjeka, dok su se linije gorila, čimpanza i bonoboa odvojile nešto kasnije. Vrijeme divergencije između ljudi i čimpanza procijenjeno je na oko 4 do 8 milijuna godina, za vrijeme kasnog miocena.[3][53][54]
Genetski su se dokazi koristili kako bi se riješilo pitanje je li postojao ikakav genski protok između ranih modernih ljudi i neandertalaca, kao i za rasvjetljavanje dinamike prvih ljudskih migracija i vremena razdvajanja različitih populacija. Uspoređujući dijelove genoma koji nisu pod pritiskom prirodnog odabira, te stoga akumuliraju mutacije jednakim tempom, moguće je rekonstruirati genetsko stablo svih ljudskih vrsta sve do posljednjeg zajedničkog pretka.
Haplogrupa se formira svaki put kada se određena mutacija (polimorfizam pojedinačnog nukleotida) pojavi u genomu pojedinca i prenese na njegovo potomstvo. Uspoređujući mitohondrijski DNK koji je naslijeđen samo od majke, genetičari su zaključili da je posljednji zajednički ženski predak svih modernih ljudi, tzv. mitohondrijska Eva, živjela prije oko 200 000 godina.
Humana evolucijska genetika proučava razlike između dva ili više ljudskih genoma, evolucijsku prošlost koja je odgovorna za ove razlike, i sadašnje učinke. Razlike među genomima imaju anthropološke, medicinske i forenzične implikacije i uporabu. Prikupljeni genetski podaci mogu nam dati važan uvid u evoluciju čovjeka.


Postoji malo fosilnih dokaza divregencije evolucijskih linija gorila, čimpanza i čovjeka.[55] Najstariji fosili koji su klasificirani kao pripadajući rodu Homo jesu ostaci Sahelanthropusa tchadensisa procijenjeni na 7 milijuna godina starosti, ostaci Orrorina tugenensisa stari oko 5,7 milijuna godina i Ardipithecusa kadabbe od oko 5,6 milijuna godina starosti. Za svakoga se od njih smatra da su bili dvonožni preci kasnijih hominina, ali su u svakom od ovih slučajeva iznesene kritike. Moguće je da je jedna ili više spomenutih vrsta predak neke druge grane afričkih majmuna ili da predstavljaju zajedničkog pretka hominina i ostalih majmuna.
Pitanje povezanosti između ovih prvih fosilnih vrsta i evolutivne linije hominina još se uvijek mora rasvijetliti. Od tih prvih vrsta odvojio se rod australopiteka prije oko 4 milijuna godina koji se razvio u dvije nama poznate grane: krupne australopiteke, poznate i kao rod Paranthropus i gracilne australopiteke. Jedna vrsta ovih posljednjih, vjerojatno Australopithecus garhi, bio je predak roda Homo. Rod australopiteka je vrlo dobro pokriven fosilnim dokazima, s vrstom Australopithecus afarensis od koje su pronađeni fosilni ostaci više od stotine jedinki kako u sjevernoj Etiopiji, tako i u Keniji i Južnoj Africi. Fosili krupnih australopiteka kao npr. Australopithecusa robustusa i Australopithecusa boisei posebno su zastupljeni u Južnoafričkoj Repubilici na lokalitetima Kromdraai i Swartkrans, te oko jezera Turkana u Keniji.
Najstariji pripadnik roda Homo je Homo habilis koji se razvio prije oko 2,3 milijuna godina. Homo habilis je prva vrsta o kojoj imamo izravne dokaze o korištenju kamenog oruđa. Pripadnici ove vrste razvili su olduvansku tehnologiju izrade kamenog oruđa, tako nazvanu po klancu Olduvai, prvom nalazištu fosila Homo habilisa. Neki znanstvenici smatraju da je Homo rudolfensis odvojena vrsta od Homo habilisa, ali većina znanstvenika smatra da su razlike među njima minorne i da se isti može klasificirati kao varijacija unutar same vrste Homo habilis, dok neki čak objašnjavaju razlike spolnim dimorfizmom. Mozak ovih prvih hominina bio je otprilike istog volumena kao u čimpanze, a njihova je osnovna karakteristika bila dvonožno hodanje i prilagodba životu na tlu.
U sljedećem milijunu godina započeo je proces encefalizacije i fosilni ostaci dokazuju da se s pojavom Homo erectusa volumen lubanje udvostručio. Homo erectus je bio prvi pripadnik hominina koji je emigrirao izvan Afrike i osim same Afrike nastanio se i u Europi i Aziji u vremenskom razdoblju od 1,8 do 1,3 milijuna godina. Jedna populacija Homo erectusa, ponekad klasificarana kao zasebna vrsta Homo ergaster, ostala je u Africi i evoluirala u Homo sapiensa. Postoje dokazi da je to prva vrsta koja je koristila vatru i izrađivala složeno kameno oruđe.
Najstariji tranzicijski fosili od Homo ergastera/erectusa prema Arhajskom Homo sapiensu pronađeni su u Africi, kao što je npr. Homo rhodesiensis, ali slični tranzicijski oblici pronađeni su i na lokalitetu Dmanisi u Gruziji. Potomci afričkog Homo erectusa raširili su se čitavom Euroazijom prije 500 000 godina i evoluirali su u Homo antecessora, Homo heidelbergensisa i Homo neanderthalensisa. Najstariji fosili anatomski modernog čovjeka datiraju iz srednjeg paleolitika prije oko 200 000 godina kao oni iz nacionalnog parka Omo u jugozapadnoj Etiopiji. Kasniji fosili modernog čovjeka, stari oko 90 000 godina, pronađeni su u pećinskoj strukturi Es Skhul u Izraelu i Južnoj Europi.
Moderni su ljudi u svom širenju izvan Afrike došli u doticaj s drugim arhaičnijim vrstama ljudi kao neandertalcima i tzv. denisovskim ljudima, koji su vjerojatno evoluirali od populacija Homo erectusa, koji su pak napustili Afriku prije oko 2 milijuna godina. Priroda interakcija između prvih modernih ljudi i arhaičnih vrsta čovjeka bila je dugo vremena predmetom kontroverzije. Među stručnjacima se vodi rasprava jesu li moderni ljudi potisnuli druge vrste i zamijenili ih ili su vrste bile dovoljno slične da je došlo do bitnijeg križanja među njima. U tom bi slučaju stare populacije hominida imale utjecaj svojim genetskim materijalom na razvoj modernog čovjeka.[56][57]
Migracije modernih ljudi izvan Afrike zbile su se pred oko 70 000 godina i moderni su se ljudi s vremenom proširili po cijelom svijetu, zamijenivši postupno druge ljudske vrste, kako nadmetanjem za resurse, tako i međusobnom hibridizacijom. Nastanili su Euroaziju i Oceaniju pred oko 40 000 godina i obje Amerike pred oko 14 500 godina.[58]
Na donjoj galeriji možemo vidjeti, kroz prijelazne oblike, čitavu povijest razvoja čovjekovih predaka od najranijeg Sahelanthropusa tchadensisa, kao prvog u nizu hominina koje više ne smatramo čovjekolikim majmunima do kromanjonaca, koje smatramo neposrednim predhodnicima ili čak podvrstom suvremene ljudske vrste Homo sapiens.
-

-
 Fosilizirana lubanja australopiteka
Fosilizirana lubanja australopiteka -
 Facijalna rekonstrukcija australopiteka
Facijalna rekonstrukcija australopiteka -
 Fosilizirana lubanja Homo habilisa
Fosilizirana lubanja Homo habilisa -
 Fosilizirana lubanja Homo erectusa
Fosilizirana lubanja Homo erectusa -
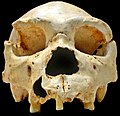 Fosilizirana lubanja Homo heidelbergensisa
Fosilizirana lubanja Homo heidelbergensisa -
 Fosilizirana lubanja Homo rhodesiensisa
Fosilizirana lubanja Homo rhodesiensisa -
 Lubanja kromanjonca
Lubanja kromanjonca -
 Digitalna facijalna rekonstrukcija kromanjonca
Digitalna facijalna rekonstrukcija kromanjonca -
 Lubanja modernog čovjeka
Lubanja modernog čovjeka -
 Portret žene masai ratnika
Portret žene masai ratnika










Evolucija povijest primata seže 65 milijuna godina natrag u vremenu.[59][60] Najstarija poznata vrsta sisavca sličnog primatima,[61] Plesiadapis, bila je raširena u Euroaziji i Africi tijekom tropskog razdoblja Paleocena i Eocena.

David Begun[62] je zaključio da rani primati izvorno potječu iz Eurazije i da je evolucijska linija koja vodi do velikih afričkih majmuna i čovjeka, uključujući Dryopithecusa, migrirala južno od Europe ili zapadne Azije u Afriku. Iz preživjele tropske populacije primata, koja je po broju fosilnih ostataka najbogatije zastupljena u fosilnim nalazištima gornjeg eocena i donjeg oligocena uvale Al Fayyum jugoistočno od Kaira, razvile su se sve živuće vrste madagaskarskih lemura i lorija iz jugoistočne Azije, afričkih galagoa ili "beba iz šipražja" i svih viših primata: majmuna Novog svijeta, catarrhinia ili majmuna Starog svijeta i velikih čovjekolikih majmuna, uključujući samog čovjeka.
Najstariji poznati ostaci pareda catarrhini jesu fosili Kamoyapithecusa iz gornjeg oligocena, pronađeni na lokalitetu Eragaleit u Velikoj rasjednoj dolini u Keniji, datirane starosti od 24 milijuna godina.[63] Pretpostavlja se da su njihovi preci bili vrste srodne Aegyptopithecusu, Propliopithecusu, i Parapithecusu iz Al Fayyuma, stari oko 35 milijuna godina.[64] Godine 2010., Saadanius je opisan kao tijesni rođak posljednjeg zajedničkog pretka pareda catarrhini, procijenjene starosti od oko 29 – 28 milijuna godina, koji je tako doprinijeo popunjavanju spoznajne rupe od 11 milijuna godina nedostataka fosilnih ostataka.[65]

U ranom miocenu, prije oko 22 milijuna godina, postojalo je u istočnoj Africi mnogo vrsta primitivnih uskonosih majmuna prilagođenih životu na stablima, koji dokazuju dugu povijest prethodne diversifikacije. Pronađeni su 20 milijuna godina stari fosilni ostaci Victoriapithecusa, najstarijeg majmuna starog svijeta. Rodovi stari 13 milijuna godina za koje smatra da spadaju u evolutivnu liniju majmuna jesu Proconsul, Rangwapithecus, Dendropithecus, Limnopithecus, Nacholapithecus, Equatorius, Nyanzapithecus, Afropithecus, Heliopithecus i Kenyapithecus, svi iz istočne Afrike.
Prisutnost drugih necercopiteka iz udaljenih fosilnih nalazišta srednjeg miocena kao Otavipithecus) iz pećina u Namibiji, Pierolapithecusa i Dryopithecusa iz Francuske, Španjolske i Austrije svjedoče o velikoj raznolikosti vrsta u Africi i mediteranskom bazenu tijekom relativno toplih i pogodnih klimatskih uvjeta ranog i srednjeg miocena. Najrecentniji čovjekoliki majmuni miocenskog razdoblja, Oreopithecusi, pronađeni su u nalazištima ugljena u Italiji i datirani su na 9 milijuna godina starosti.
Dokazi iz molekularne biologije pokazuju da se evolucijska grana gibona (porodica Hylobatidae) odvojila od čovjekolikih majmuna prije oko 18 do 12 milijuna godina i da se grana orangutanovih predaka (potporodica Ponginae) odvojila prije 12 milijuna godina. Ne postoje fosilni ostaci koji bi jasno dokumentirali pretke gibona, koji su se mogli razviti iz još uvijek nepoznate populacije majmuna iz jugoistočne Azije, ali fosili protoorangutana su jasno dokumentirani ostacima Sivapithecusa iz Indije i Griphopithecusa iz Turske, datirane starosti od oko 10 milijuna godina.[66]
Vrste bliske posljednjem zajedničkom pretku gorila, čimpanza i ljudi su fosili Nakalipithecusa otkriveni u Keniji i Ouranopithecusa pronađeni u Grčkoj. Dokazi iz molekularne biologije navode da su se preci prvih gorila, potom čimpanza (rod Pan) odvojili od evolucijske linije koja vodi ka čovjeku prije 8 do 4 milijuna godina. Ljudska DNK je otprilike 98.4% jednaka onoj u čimpanze, ako se analiziraju polimorfizmi pojedinačnog nukleotida. Otkriveni fosilni ostaci predaka gorila i čimpanza su rijetki. Loše očuvanje fosilnih ostataka u tropskim područjima i mali broj fosila mogu nam dati iskrivljenu sliku evolutivnog razvoja ovih dvaju rodova.
Druge su se vrste hominina prilagodile suhom okolišu izvan ekvatorijalnog pojasa zajedno s faunom antilopa, hijena, pasa, svinja, slonova i konja. Ekvatorijalni pojas se suzio prije oko 8 milijuna godina. Postoji malo fosilnih dokaza odvajanja evolutivne linije hominina od linija gorila i čimpanza. Najstariji fosili za koje se smatra da pripadaju evolutivnoj liniji čovjeka jesu Sahelanthropus tchadensis (star 7 milijuna godina) i Orrorin tugenensis (star 6 milijuna godina), zatim slijedi rod Ardipithecus (star 5.5–4.4 milijuna godina), s poznatim vrstama Ardipithecus kadabba i Ardipithecus ramidus;
Rod Australopithecus je evoluirao u istočnoj Africi prije oko 4 milijuna godina i proširio se po cijelom afričkom kontinentu, a izumro je prije 2 milijuna godina. U tom je vremenskom razdoblju postojalo više vrsta australopiteka, od kojih su nama poznati Australopithecus anamensis, Australopithecus afarensis, Australopithecus sediba i Australopithecus africanus. Stručnjaci još uvijek imaju dvojbe da li uvrstiti neke vrste afričkih hominida,kao što je npr. Australopithecus robustus i Australopithecus boisei u isti rod australopiteka ili u zaseban rod. Ako ih se klasificira kao pripadnike australopiteka, tada ih se mora smatrati podskupinom "krupnih australopiteka", dok se ostali smatraju "gracilim australopitecima".
Slijedi popis vrsta australopiteka:
- Australopiteci (od 4 do 1.8 milijuna godina starosti) s vrstama Australopithecus anamensis, Australopithecus afarensis, Australopithecus africanus, Australopithecus bahrelghazali, Australopithecus garhi i Australopithecus sediba;
- Kenyanthropus (od 3 do 2.7 milijuna godina starosti) s vrstom Kenyanthropus platyops;
- Paranthropus (od 3 do 1.2 milijuna godina starosti) s vrstama Paranthropus aethiopicus, Paranthropus boisei iParanthropus robustus.

Homo sapiens je jedina preživjela vrsta roda Homo. Iako neki pronađeni fosili pripadaju izumrlim vrstama roda Homo koji su vjerojatno izravni preci Homo sapiensa, većina su najvjerojatnije bočne grane čovjekove evolucijske linije.[68][69] Ne postoji konsenzus koja se od ovih izumrlih vrsta može smatrati posebnom vrstom, a koja podvrstom. Razlog tome je u nekim slučajevima oskudnost fosilnih ostataka, dok je u drugim slučajevima velika anatomska sličnost pojedinih vrsta roda Homo.[69] Teorija prema kojoj je postojalo razdoblje obilnih padalina na području današnje Sahare daje nam moguće objašnjenje rane raznolikosti i lakoće migracija unutar roda Homo.
Na temelju arheoloških i paleontoloških dokaza, moguće je do neke mjere spoznati dijetalne navike različitih vrsta roda Homo i proučiti utjecaj prehrane na fizičku evoluciju i evoluciju u ponašanju unutar samog roda.[70][71][72][73][74] Mnogi antropolozi smatraju da je erupcija supervulkana Toba na otoku Sumatri u Indoneziji prije oko 70 000 godina, imala globalne klimatske posljedice[75] i usmrtila većinu ljudi onog doba, stvoreći tako pupulacijsko usko grlo koje je smanjilo genetsku raznolikost većine današnjih ljudi.[76]
Homo habilis je živio prije 2,3 do 1,65 milijuna godina. Evoluirao je u južnoj i istočnoj Africi u kasnom pliocenu i ranom pleistocenu prije oko 2,5 do 2 milijuna godina, odvojivši se od evolutivne grane australopiteka. Homo habilis je imao manje kutnjake i veći mozak od australopiteka i izrađivao je kameno oruđe i vjerojatno oruđe od kosti. Jednome od prvih poznatih hominida otkrivenih od "lovca na fosile" Louisa Leakeya nadjenut je nadimak "spretni čovjek" (eng.: handy man) zbog njegove sposobnosti korištenja kamenog oruđa. Neki su znanstvenici predložili da se ova vrsta premjesti iz roda Homo u rod Australopithecusa zbog morfologije kostura koji je bolje prilagođen životu na stablu nego dvonožnom hodanju.[77]
Homo habilis se smatrao prvom poznatom vrstom roda Homo, sve do svibnja 2010. kad je u južnoj Africi otkrivena nova vrsta Homo gautengensis, kronološki nešto starija od Homo habilisa, iako je valjanost te vrste osporena.[78]
Homo rudolfensis i Homo georgicus su imena dviju vrsta, čiji su fosilni ostaci stari od 1,9 do 1,6 milijuna godina. Njihova veza s Homo habilisom je još nejasna.
Homo rudolfensis je ime za vrstu čiji je jedini poznati ostatak jedna nepotpuna lubanja pronađena u Keniji. Znanstvenici su predložili da je to varijanta Homo habilisa, ali to još uvijek nije potvrđeno.[79] Homo georgicus, čiji su fosilni ostaci otkriveni u Gruziji, vjerojatno je prijelazni oblik između Homo habilisa i Homo erectusa,[80] ili podvrsta Homo erectusa.[81]
Prve fosile Homo erectusa otkrio je nizozemski liječnik Eugene Dubois 1891. na indonezijskom otoku Javi. On je ovu vrstu u početku nazvao Pithecanthropus erectus na temelju njegove morfologije i smatrao ga je posrednom vrstom majmuna i čovjeka.[82] Fosilni ostaci Homo erectusa pronađeni na Javi danas su poznati i kao Javanski čovjek. Homo erectus je živio u vremenskom razdoblju od prije 2 milijuna do 100 000 godina, a Homo erectus soloensis na Javi je posljednja podvrsta koja je izumrla. Rani ostaci ove vrste, od 1,8 do 1,25 milijuna godina, često se klasificiraju kao ostaci odvojene vrste Homo ergastera, ili se smatraju podvrstom Homo erectusa, (Homo erectus ergaster).
U ranom pleistocenu, od 1,5 do 1 milijun godina, neke su populacije Homo habilisa u Africi evoluirale razvijajući veći volumen mozga, uspravno držanje, te su počele izrađivati elaborirano kameno oruđe. Ove su razlike između ostalog dovoljne da ih antropolozi klasificiraju kao novu vrstu, Homo erectusa.[83] Uspravno je držanje postalo moguće evolucijom koljena koje se potpuno moglo izravnati i pozicioniranjem foramen magnuma sa stražnjeg u donji dio lubanje. Homo erectus je znao koristiti vatru za termičku obradu mesa.
Fosilni ostaci Homo erectusa pronađeni su diljem Azije, Afrike i Europe, od kojih je jedan od napoznatijih Pekinški čovjek. Mnogi paleoantropolozi koriste naziv Homo ergaster za neazijske oblike ove vrste, a nazivaju Homo erectusom samo one fosile pronađene u Aziji, temeljeći ovu podjelu na manjim razlikama u građi kostiju i zubi.
Homo antecessor je vrsta čiji su ostaci pronađeni u Španjolskoj i Engleskoj i njihova je starost datirana od 1,2 milijuna do 800 000 godina. DNK analiza je pokazala da je ova vrsta zaseban evolucijski ogranak od Homo erectusa[84][85]
Homo cepranensis je moguća vrsta od koje je pronađen samo dio lubanje u Italiji, koja je procijenjena na 800 000 godina starosti. Većina paleontologa smatra da lubanja pripada ili Homo erectusu ili Homo heildebergensisu.[86]
Homo heidelbergensis (ili čovjek iz Heidelberga) je vrsta koja je živjela prije 800 000 do prije 300 000 godina. Predloženi su i alternativni nazivi Homo sapiens heidelbergensis ili Homo sapiens paleohungaricus.[87]
Homo rhodesiensis (ili čovjek iz Rodezije) je vrsta čija se starost procjenjuje na 300 000 do 125 000 godina. Većina današnjih znanstvenika klasificiraju ovu vrstu u istu grupu s Homo heidelbergensisom, iako su predložene i alternativne klasifikacije, ili kao zasebna vrsta ili kao arhajski Homo sapiens.
U veljači 2006. u Etiopiji je pronađen fosil, danas poznat kao lubanja iz Gawisa, koji bi mogao biti međuvrsta između Homo erectusa i Homo sapiensa ili jedna od mnogih evolucijskih slijepih ulica. Procijenjuje se da je lubanja iz Gawisa stara od 500 000 do 250 000 godina. Poznto je vrlo malo detalja o ovm pronalasku, pošto još nije objavljena studija u stručnoj literaturi.[88]
Homo neanderthalensis ili Homo sapiens neanderthalensis,[89] je živio u Europi i Aziji prije 400 000 sve do prije 30 000 godina. Prve genetske analize mitohondrijske DNK pokazale su da nije bilo bitnijeg genskog protoka između Homo neanderthalensisa i Homo sapiensa, one se dakle smatraju odvojenim vrstama čiji je posljednji zajednički predak živio prije 660 000 godina.[90][91][92] Međutim, 2010. godine dovršen je ambiciozni projekt potpunog sekvenciranja genoma neandertalaca, koji je dokazao da su se neandertalci uistinu križali s anatomski modernim ljudima prije oko 45 000 do 80 000 godina (što se podudara s razdobljem napuštanja Afrike od strane modernih ljudi i njihovog širenja Europom i Azijom.[93]
Dokazano je da 1% do 4% DNK gotovo svih modernih ljudi koji nisu afričkog podrijetla derivira iz DNK neandertalaca,[93] a ovo je otkriće sukladno nedavnim studijama koje dokazuju da se divergencija nekih ljudskih alela zbila prije oko milijun godina, iako takva interpretacija ovih rezultata može biti dovedena u pitanje.[94][95] Konkuriranje za iste resurse s Homo sapiensom vjerojatno je dovelo do izumiranja neandrtalaca.[96][97] Ove su vrste koegzistirale u Europi oko 10 000 godina, tijekom kojih je populacija modernih ljudi naglo narasla u odnosu na neandertalce i vjerojatno ih potisnula samim brojem jedinki.[98]
Godine 2008., skupina arheologa koji su istraživali špilja Denisova na gorju Altaj u Sibiru otkrili su mali komad kosti prsta mlade jedinke grupe hominina danas poznatog kao denisovski čovjek.[99] Pronađeni artefakti, uključujući i jednu narukvicu, nalazili su se u istom geološkom sloju s kostima i radiodatirani su ugljikom 14 pokazujući starost od oko 40 000 godina. Pošto se DNK dobro očuvao zahvaljujući i vrlo hladnim klimatskim uvjetima u samoj pećini, bilo je moguće izolirati i sekvencirati DNK cijelog genoma.[10][100]
Iako je analiza mtDNK pokazala da je vrijeme razilažnja evolucijskih linija denisovskog čovjeka i modernog čovjeka neočekivano pomaknuta unatrag u vremenu,[101] sekvenciranje cijelog genoma dokazuje da denisovski ljudi pripadaju istoj evolucijskoj liniji kao i neandertalci i da su se njihove evolucijske linije odvojile nedugo nakon njihovog odvajanja od evolucijske linije modernih ljudi.[10] Poznato je da su moderni ljudi živjeli u isto vrijeme s neandertalcima više od 10 000 godina, a otkriće denisovskog čovjeka otvara mogućnost da su neandertalci, denisovski ljudi i moderni ljudi koegzistirali i interagirali. Koegzistencija tako različitih vrsta hominina ukazuje na puno složeniju sliku čovječanstva tijekom kasnog pleistocena nego što se prije mislilo.[100][102] Dokazano je da oko 6% genoma modernih stanovnika Melanezije vuće podrijetlo od denisovskiog čovjeka, dokazujući ograničeno križanje dviju vrsta u Jugoistočnoj Aziji.[103][104]
Aleli za koje postoje naznake da pripadaju neandertalcima i denisovskim ljudima pronađeni su u nekoliko genskih lokusa modernih ljudi izvan Afrike. HLA aleli neandertalaca i denisovaca predstavljaju polovicu HLA alela modernih ljudi Europe i Azije,[46] ukazujući na jaku pozitivnu selekciju za ove alele koji proizlaze iz međusobnog križanja različitih vrsta.
Homo floresiensis je živio u vremenskom razdoblju od prije 190 000 do prije 50 000 godina. Dobio je nadimak hobbit zbog njegove male tjelesne građe, vjerojatno prouzročene procesom otočne patuljastosti.[105] Homo floresiensis je intrigantan slučaj za stručnjake kako zbog veličine tijela tako i zbog relativno male starosti, pošto je to recentna vrsta roda Homo koja pokazuje vrlo različite tjelesne karakteristike u odnosu na moderne ljude. Drugim riječima, Homo floresiensis dijeli zajedničkog pretka s modernim ljudima, ali se u prošlosti odvojio od evolutivne linije modernih ljudi i krenuo posebnom evolucijskom linijom.
Glavno otkriće ove vrste je kostur žene stare oko 30 godina pronađen 2003. datirane starosti od oko 18 000 godina. Žena kojoj su pripadali fosilni ostaci bila je visoka oko jedan metar i imala je moždani volumen od samo 380 cm3. Ovakav se moždani volumen smatra malim i za čimpanzu i predstavlja jednu trećinu moždanog volumena suvremenog čovjeka.
Međutim, vodi se rasprava da li je Homo floresiensis uistinu zasebna vrsta.[106] Neki znanstvenici tvrde da je Homo floresiensis bio moderni Homo sapiens koji je bolovao od patološke nanosomije.[107] Ova je hipoteza djelomično podržana i činjenicom da su neki pripadnici domorodaca koji dan danas žive na otoku Flores pigmeji. Niski rast pigmejaca zajedno s patološkom nanosomijom mogao je kao posljedicu imati postojanje ljudi s izuzetno sitnom tjelesnom građom. Još jedna činjenica koja izravno napada hipotezu o Homo floresiensisu kao zasebnoj biološkoj vrsti je pronalazak i kamenog oruđa tipičnog samo za Homo sapiensa u istom fosilnom nalazištu.[107]
Hipoteza o patološkoj nanosomiji, međutim, ne može objasniti dodatne anatomske karakteristike netipične za modernog čovjeka (bolesnog ili ne), ali vrlo moguće kod pripadnika različite vrste roda Homo. Osim karakteristika lubanje, dodatne karakteristike netipične za modernog čovjeka koje nalazimo kod čovjeka iz Floresa su morfološke karakteristike zgloba šake, podlaktice, ramena, koljena i stopala. Osim toga, hipoteza patološke nanosomije ne objašnjava pronalazak više primjeraka jedinki sitne tjelesne građe, sugerirajući da je to bila opća karakteristika šire populacije na tom lokalitetu, a ne ograničena na jednu oboljelu jedinku.
Homo sapiens (hrv.: razumni, inteligentni čovjek) pojavio se prije oko 315 000 godina i jedina je živuća vrsta roda Homo. Između 400 000 godina i 250 000 godina, tijekom cijelog interglacijalnog razdoblja srednjeg pleistocena, trajao je evolucijski trend povećavanja moždanog volumena i razvoja tehnologije obrade kamenog oruđa, od pojave Homo erectusa sve do Homo sapiensa. Izravni dokazi ukazuju da je postojala migracija Homo erectusa izvan Afrike, a kasnijom specijacijom na afričkom kontinentu od Homo erectusa se je razvio Homo sapiens. Kasnije migracije unutar i izvan afričkog kontinenta postupno su zamijenile populacije Homo erectusa iz ranijih migracija. Ova teorija pojave modernog čovjeka na afričkom kontinentu i kasnijih migracija poznata je kao teorija nedavnog afričkog pretka ili teorija "Out of Africa". Trenutni dokazi ne isključuju u potpunosti određeni stupanj multiregionalne evolucije i DNK analiza dokazuje miješanje Homo sapiensa s populacijama drugih vrsta roda Homo. Oko ovog se pitanja vode žestoke diskusije među paleoantropolozima.
Novija su istraživanja dokazala da su ljudi u genetskom smislu vrlo homogeni, tj. postoji vrlo mala varijabilnost u ljudskoj DNK u odnosu na druge životinjske vrste. To može biti posljedica relativno recentnog evolucijskog razvoja ljudske vrste ili pak posljedica populacijskog uskog grla koji je nastao zbog kataklizmičkih događaja kao erupcije supervulkana Tobe.[108][109][110] Posebne genetske karakteristike su se, međutim, razvile kao rezultat seoba malih ljudskih skupina u nova područja i njihove prilagodbe novim okolišnim uvjetima. Ove nove genetske karakteristike čine mali dio genoma Homo sapiensa i obuhvaćaju boju kože, oblik nosa i mogućnost učinkovitijeg disanja na velikim nadmorskim visinama.
Homo sapiens idaltu, čiji su fosilni ostaci pronađeni u Etiopiji, je izumrla podvrsta Homo sapiensa od prije 160 000 godina.
| vrsta | vrijeme življenja (milijuni godina) | mjesto življenja | visina odrasle jedinke | težina odrasle jedinke | volumen lubanje (neurocraniuma) (cm³) | fosilni dokazi | otkriće / publikacija imena |
|---|---|---|---|---|---|---|---|
| denisovski čovjek | 0.04 | Rusija | 1 lokalitet | 2010. | |||
| Homo antecessor | 1.2 – 0.8 | Španjolska | 175 cm | 90 kg | 1 000 | 2 lokaliteta | 1997. |
| Homo cepranensis (upitna valjanost vrste) | 0.9 – 0.35 | Italija | 1 000 | dio lubanje | 1994./2003. | ||
| Homo erectus | 2 – 0.1 | Afrika, Eurazija (Java, Kina, Indija, Kavkaz) | 180 cm | 60 kg | 850 (rani) – 1 100 (kasni) | mnogi | 1891./1892. |
| Homo ergaster (ili Homo erectus ergaster) | 1.9 – 1.4 | istočna i južna Afrika | 190 cm | 700–850 | mnogi | 1975. | |
| Homo floresiensis | 0.19 – 0.05 | Indonezija | 100 cm | 25 kg | 400 | 7 jedinki | 2003./2004. |
| Homo gautengensis (upitna valjanost vrste) | >2 – 0.6 | južna Afrika | 100 cm | 1 jedinka | 2010./2010. | ||
| Homo habilis | 2.3 – 1.65 | Afrika | 150 cm | 33–55 kg | 510–660 | mnogi | 1960./1964. |
| Homo heidelbergensis | 0.6 – 0.35 | Europa, Afrika, Kina | 180 cm | 90 kg | 1 100–1 400 | mnogi | 1908. |
| Homo neanderthalensis | 0.35 – 0.04 | Europa, zapadna Azija | 170 cm | 55 – 70 kg (robustne građe) | 1 200–1 900 | mnogi | (1829.)/1864. |
| Homo rhodesiensis (upitna valjanost vrste) | 0.3 | Zambija | 1 300 | vrlo malo | 1921. | ||
| Homo rudolfensis | 1.9 - 1.5 | Kenija | 700 | 2 lokaliteta | 1972./1986. | ||
| Homo naledi | 0.3 - 0.2 | Južna Afrika | mnogi | 2013. | |||
| Homo sapiens idaltu | 0.16 – 0.15 | Etiopija | 1 450 | 3 lubanje | 1997./2003. | ||
| Homo sapiens (moderni čovjek) |
0.3 – danas | cijeli svijet | 150 – 190 cm | 50–100 kg | 950–1 800 | živuća vrsta | — /1758. |
 |




Izrada alata se interpretira kao znak inteligencije, i pretpostavlja se da je uporaba alata mogla stimulirati određene aspekte ljudske evolucije, pogotovo stalnog povećanja ljudskog mozga. Paleontolozi još uvijek proučavaju sve aspekte ove tendencije povećananja ljudskog mozga tijekom evolutivnog razvoja, unatoč činjenici da je mozak vrlo zahtjevan u smislu utroška energije. Mozak modernog čovjeka troši oko 13 watti dnevno (oko 260 kilokalorija dnevno), što je jedna petina ukupnog dnevnog utroška energije.[111]
Povećana uporaba alata omogućuje lov na energetski bogate izvore mesa i obradu energetski bogatih izvora iz biljaka. Znanstvenici tumače da su rani hominidi stoga bili pod jakim evolutivnim pritiskom u smjeru povećanja sposobnosti uporabe alata.[112] Vrijeme početka uporabe alata od strane prvih ljudi je teško odrediti, jednostavno stoga što je vrlo teško odrediti da li je vrlo primitivno oruđe (npr. kamenje oštrih rubova) prirodni predmet ili ljudski artefakt. Postoje određeni dokazi da su australopiteci prije 4 milijuna godina koristili krhotine kostiju kao alat, ali to je još uvijek predmet diskusija.[113]
Mora se uzeti u obzir da mnoge životinjske vrste izrađuju i koriste alat, ali su samo vrste roda Homo uspjele ovladati životnim okolišem izradom i uporabom složenijeg oruđa. Najstariji poznati alat je "oldovansko kameno oruđe" iz Etiopije procijenjene starosti od 2,5-2,6 milijuna godina. Ne postoje poznati dokazi postojanja bilo kakvog ljudskog artefakta starijeg od 2,5 milijuna godina. Fosil roda Homo pronađen je u blizini oldovanskog oruđa, i njegova je starost procijenjena na 2,3 milijuna godina, ukazujući na to da su pripadnici hominina uistinu izrađivali i koristili kameno oruđe. Ova je hipoteza najvjerojatnija, ali još uvijek ne predstavlja čvrsti dokaz.[114] Razvoj stiloidnog procesa treće metakarpalne kosti omogućio je kostima šake zatvaranje unutar kostiju zgloba šake, omogućujući jači stisak same šake. To je omogućilo ljudima snagu i spretnost korištenja složenog oruđa. Ova jedinstvena anatomska karakteristika razlikuje ljude od suvremenih majmuna i neljudskih primata, i nije prisutna u ljudskim ostacima starijim od 1,8 milijuna godina.[115]
Bernard Wood je primijetio da su vrste roda Paranthropus koegzistirale s ranim vrstama roda Homo na istom području gdje je pronađeno oldovansko kameno oruđe okvirno u istom vremenskom razdoblju. Iako ne postoje izravni dokazi da su pripadnici Paranthropusa izrađivali kameno oruđe, njihova anatomija neizravno ukazuje na njihove mogućnosti izrade. Većina se paleoantropologa slaže da su rane vrste ljudi pravi stvaratelji većine oldovanskog kamenog oruđa. Oni navode da je oldovansko oruđe uvijek bilo prisutno u bilizini fosilnih ostataka vrsta Homo, ali ne i u blizini vrsta Paranthropus.[114]
Godine 1994. Randall Susman je postulirao postojanje anatomske karakteristike odvojenog palca kao argument da su i Homo i Paranthropus vrste izrađivale oruđe. On je usporedio mišiće palca čovjeka i čimpanze, i otkrio da ljudi imaju razvijena tri mišića koji nedostaju kod čimpanzi. Ljudi imaju i deblje kosti zapešća sa širim glavama, omogućujući tako prciznije hvatanje predmeta nego to mogu učiniti čimpanze. Susman pretpostavlja da je moderna anatomija ljudskog palca evolucijski odgovor na povećane potrebe manipuliranja predmetima vezane za izradu i korištenje kamenog oruđa.[114]
Najstarije dokazano kameno oruđe staro je 2,6 milijuna godina, kad je Homo habilis u istočnoj Africi koristio tzv. oldovansko oruđe od kamenih oblutaka. Izrađivao je krhotine oštrih rubova lomeći glatke oblutke jednostavnim udarcima kamen o kamen.[116] Ovo se razdoblje označava kao početak paleolitika ili starijeg kamenog doba, koje završava s krajem posljednjeg ledenog doba prije 10 000 godina. Paleolitik se dijeli na donji paleolitik (rano kameno doba, koje završava razdobljem prije 350 000–300 000 godina), srednji paleolitik (srednje kameno doba, do prije 50 000–30 000 godina) i gornji paleolitik.
Razdoblje od prije 700 000 do 300 000 godina poznato je kao ašelejenska kultura, u kojem je Homo ergaster (ili Homo erectus) izrađivao široke ručne sjekire od kremena i kvarcita, u početku prilično grube izrade (rana ašelejeska kultura), a kasnije dodatno obrađene odlamanjem sitnih krhotima finim udarcima po rubovima kamenog instrumenta. Oko 350 000 godina prije današnjice pojavio se rafiniraniji način obrade poznat kao levaloaška tehnika, koja se zasniva na prethodnoj pripremi kamene jezgre radi dobivanja odbitka standardne forme i veličine. Ovakav odbitak se kasnije ponovo obrađivao (sekundarna obrada), do konačnog oblika, najčešće rezača, šiljaka i strugača.[116] Na posljetku, otprilike 50 000 godina prije današnjice, pojavilo se još više rafinirano i specijalizirano kremeno oruđe u populacijama neandertalaca i doseljenih kromanjonaca (kameni noževi, oštrice, strugači). U tom su razdoblju započeli izradu i prvog oruđa od kostiju.
Do prije 50 000 – 40 000 tisuća godina uporaba kamenog oruđa se polako i postupno razvijala. Svaka bi faza (Homo habilis, Homo ergaster, Homo neanderthalensis) krenula s više razine složenosti od prethodne, ali nakon što se svaka faza pokrenula dalji je razvoj bio spor. Paleoantropolozi trenutno vode rasprave da li su ove vrste roda Homo imale neke od mnogih kulturalnih karakteristika i karakteristika u ponašanju tipične za moderne ljude, kao npr. jezik, složeno simboličko razmišljanje i tehnološku kreativnost. Prethodne su vrste roda Homo naizgled bile kulturalno konzervativne, te su se držale jednostavnih tehnologija i jednakih obrazaca potrage za hranom kroz vrlo duga vremenska razdoblja.
Oko 50 000 godina prije današnjice, čovjekova se kultura počela puno brže razvijati. Tranzicija k modernosti u ljudskom ponašanju definirana je kao eurazijski "veliki skok naprijed",[117] ili kao "revolucija gornjeg paleolitika",[118] zbog iznenadne pojave karakterističnih znakova modernog ponašanja u arheološkim nalazima. Neki znanstvenici pretpostavljaju da je ovaj prijelaz bio postupniji, jer su se neke značajke pojavile već kod arhajskog afričkog Homo sapiensa prije oko 200 000 godina.[119][120]
Moderni su ljudi počeli pokapati mrtve, koristiti životinjsko krzno kao odjeću, koristiti sofisticirane tehnike lova kao npr. rupe u terenu kao zamke ili tjeranje velikih životinja u provalije i prikazivati događaje iz svog života kroz spiljsko slikarstvo.[121] S razvojem kulture, različite su populacije unosile novosti u postojeće tehnologije: artefakti poput udica, dugmadi i igala izrađenih od kostiju pokazivali su značajne varijacije među različitim ljudskim populacijama, a tako se nešto nikad nije desilo u ljudskim kulturama u razdobljima prije 50 000 godina od današnjice. Karakterističan primjer su različite populacije neandertalaca koje su sve imale jednaku tehnologiju i kulturu, iako su bile raširene na velikom geografskom području.
Među specifične primjere modernog ljudskog ponašanja, antropolozi ubrajaju korištenje nakita, spiljsko slikarstvo, organiziranje životnog prostora, postojanje složenih rituala (pokapanje mrtvih s poklonima položenim u grob), specijalizirane tehnike lova, istraživanje negostoljubivih područja, i mreže trgovanja trampom. Još uvijek se vodi rasprava da li se je zbila relativno brza "revolucija" koja je dovela do modernog ponašanja (tzv. "veliki prasak ljudske svjesnosti"), ili je proces bio postupan.[122]
Prirodni odabir se i danas zbiva među ljudskim populacijama. Populacije koje su, na primjer, pod velikim rizikom oboljevanja od teške prionske bolesti kuru imaju značajniju ekspresiju imune varijante gena prionskog proteina G127 V u odnosu na neimune alele. Frekvencija ove mutirane varijante je posljedica preživljavanja pojedinaca nositelja mutacije.[123] Drugi poznati evolutivni trendovi su produljenje reprodukcijskog perioda, smanjenje razina kolesterola i glukoze u krvi, te smanjenje krvnog tlaka.
Znanstvenici tvrde da se evolucija čovjeka ubrzala od kad je čovjek ovladao poljoprivredom i razvio civilizaciju prije 10 000 godina. Tvrde da je to rezultiralo značajnim genetskim razlikama među različitim postojećim ljudskim populacijama.[124] Očuvanje ekspresije enzima laktaze primjer je nedavne evolucije. Nedavna ljudska evolucija je, međutim, uglavnom ograničena na razvijanje genske rezistencije na infektivne bolesti, koje su se pojavile među ljudima kao posljedica blizine s domaćim životinjama.[125] Povećanje međusobnog križanja među populacijama ljudi od početka poljoprivrede smanjio je genski drift i geografsku izolaciju.
S druge strane, mnogi pak tvrde da je razvoj poljoprivrede usporio, ako ne i zaustavio evoluciju čovjeka, pošto postoje solidni dokazi da su rani moderni ljudi kao kromanjonci bili jači i tjelesno razvijeniji od ljudi sadašnjice.[126][127]
Slijedi popis vrsta po vremenskom redoslijedu podijeljenih po rodu:
|
|
- ↑ Heng HH. Svibanj 2009. The genome-centric concept: resynthesis of evolutionary theory. BioEssays. 31 (5): 512.–525. doi:10.1002/bies.200800182. ISSN 0265-9247. PMID 19334004
- ↑ "Nova – Meet Your Ancestors". PBS.
- ↑ a b Dawkins R. (2004.) "The Ancestor's Tale". Time Tree. 2009.
- ↑ Java Man, Curtis, Swisher and Lewin, ISBN 0-349-11473-0
- ↑ Stringer, C.B. 1994. Evolution of Early Humans. Jones, Steve; Martin, Robert; Pilbeam, David (ur.). The Cambridge Encyclopedia of Human Evolution. Cambridge University Press. Cambridge. str. 242. ISBN 978-0-521-32370-3
- ↑ McHenry, H.M. 2009. Human Evolution. Ruse, Michael; Travis, Joseph (ur.). Evolution: The First Four Billion Years. The Belknap Press of Harvard University Press. Cambridge, Massachusetts. str. 265. ISBN 978-0-674-03175-3
- ↑ Out of Africa Revisited - 308 (5724): 921g - Science. Sciencemag.org. 13. svibnja 2005. doi:10.1126/science.308.5724.921g. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 23. studenoga 2009.
- ↑ Nature. 12. lipnja 2009. Human evolution: Out of Ethiopia. Nature. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 23. studenoga 2009.
- ↑ Origins of Modern Humans: Multiregional or Out of Africa?. ActionBioscience. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 23. studenoga 2009.
- ↑ a b c Reich D., Green R.E., Kircher M.; i dr. Prosinac 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature. 468 (7327): 1053.–1060. Bibcode:2010Natur.468.1053R. doi:10.1038/nature09710. PMID 21179161 Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: više imena: authors list (link) - ↑ Noonan J. P. Srpanj 2010. Neanderthal genomics and the evolution of modern humans. Genome Res. 20 (5): 547.–553. doi:10.1101/gr.076000.108076000.108. PMC 2860157. PMID 20439435
- ↑ a b Laurent Abi-Rached; i dr. 25. srpnja 2011. The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans. Science. 334 (6052). Bibcode:2011Sci...334...89A. doi:10.1126/science.1209202. PMID 21868630. Inačica izvorne stranice arhivirana 15. travnja 2023. Pristupljeno 2. rujna 2012. Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: bot: nepoznat status originalnog URL-a (link) - ↑ Fossil Reanalysis Pushes Back Origin of Homo sapiens, 17. veljače 2005., Scientific American.
- ↑ Mellars, Paul (2006.). "Why did modern human populations disperse from Africa ca. 60,000 years ago?". Proceedings of the National Academy of Sciences 103 (25): str. 9381.–9386. Bibcode 2006PNAS..103.9381M. DOI:10.1073/pnas.0510792103 . PMC 1480416. PMID 16772383.
- ↑ Boyd, Robert; Silk, Joan B. 2003. How Humans Evolved. Norton. New York, New York. ISBN 0-393-97854-0CS1 održavanje: više imena: authors list (link)
- ↑ Brues, Alice M.; Snow, Clyde C. 1965. Physical Anthropology. Biennial Review of Anthropology. 4: 1.–39.CS1 održavanje: više imena: authors list (link)
- ↑ Brunet, M.; Guy, F.; Pilbeam, D.; Mackaye, H.; Likius, A.; Ahounta, D.; Beauvilain, A.; Blondel, C.; Bocherens, H.; Boisserie, J.; De Bonis, L.; Coppens, Y.; Dejax, J.; Denys, C.; Duringer, P.; Eisenmann, V.; Fanone, G.; Fronty, P.; Geraads, D.; Lehmann, T.; Lihoreau, F.; Louchart, A.; Mahamat, A.; Merceron, G.; Mouchelin, G.; Otero, O.; Pelaez Campomanes, P.; Ponce De Leon, M.; Rage, J.; Sapanet, M.; Schuster, M.; Sudre, J.; Tassy, P.; Valentin, X.; Vignaud, P.; Viriot, L.; Zazzo, A.; Zollikofer, C. 2002. A new hominid from the Upper Miocene of Chad, Central Africa. Nature. 418 (6894): 145.–151. doi:10.1038/nature00879. PMID 12110880CS1 održavanje: više imena: authors list (link)
- ↑ a b P. Thomas Schoenemann. 2006. Evolution of the Size and Functional Areas of the Human Brain. Annu. Rev. Anthropol. 35: 379.–406.
- ↑ Park, Min S.; Nguyen, Andrew D.; Aryan, Henry E.; U, Hoi Sang; Levy, Michael L.; Semendeferi, Katerina. 2007. Evolution of the human brain: changing brain size and the fossil record. Neurosurgery. 60 (3): 555.–562. doi:10.1227/01.NEU.0000249284.54137.32. PMID 17327801CS1 održavanje: više imena: authors list (link)
- ↑ Bruner, Emiliano. 2007. Cranial shape and size variation in human evolution: structural and functional perspectives (PDF). Child's Nervous System. 23 (12): 1357.–1365. doi:10.1007/s00381-007-0434-2. PMID 17680251. Inačica izvorne stranice (PDF) arhivirana 2. ožujka 2012. Pristupljeno 2. rujna 2012.
- ↑ Potts, Richard. 2012. Evolution and Environmental Change in Early Human Prehistory. Annu. Rev. Anthropol. 41: str. 151.–167.
- ↑ Leonard, William R., J. Josh Snodgrass, and Marcia L. Robertson. 2007. Effects of Brain Evolution on Human Nutrition and Metabolism. Annu. Rev. Nutr. 27: str. 311.–327.
- ↑ Meat-eating was essential for human evolution, says UC Berkeley anthropologist specializing in diet. Berkeley.edu. 14. lipnja 1999. Pristupljeno 31. siječnja 2012.
- ↑ Meat in the human diet: an anthropological perspective. - Free Online Library. Thefreelibrary.com. 1. rujna 2007. Pristupljeno 31. siječnja 2012.
- ↑ Organ, Chris. 22. kolovoza 2011. Phylogenetic rate shifts in feeding time during the evolution of Homo. PNAS. Pristupljeno 17. travnja 2012.
- ↑ Darwin, Charles. 1871. The Descent of Man, and Selection in Relation to Sex. Ovo je izdanje iz 1981., s uvodom Johna Tylera Bonnera i Roberta M. Maya. izdavač: Princeton University Press. Princeton NJ. ISBN 0-691-02369-7
- ↑ Behar et al. 2008, Gonder et al. 2007, Reed and Tishkoff
- ↑ Modern Humans Came Out of Africa, "Definitive" Study Says. News.nationalgeographic.com. 28. rujna 2010. Pristupljeno 14. srpnja 2011.
- ↑ Stringer CB, Andrews P. Ožujak 1988. Genetic and fossil evidence for the origin of modern humans. Science. 239 (4845): 1263.–1268. Bibcode:1988Sci...239.1263S. doi:10.1126/science.3125610. PMID 3125610
- ↑ Wolpoff, MH; Hawks, J; Caspari, R. 2000. Multiregional, not multiple origins. Am J Phys Anthropol. 112 (1): 129.–136. PMID 10766948
|url-status=deadzahtijeva|archive-url=(pomoć) - ↑ Wolpoff, MH; Spuhler, JN; Smith, FH; Radovcic, J; Pope, G; Frayer, DW; Eckhardt, R; Clark, G. 1988. Modern Human Origins. Science. 241 (4867): 772.–774. Bibcode:1988Sci...241..772W. doi:10.1126/science.3136545. PMID 3136545
- ↑ Amade M'charek. 2005. The Human Genome Diversity Project: an ethnography of scientific practice. Cambridge University Press. str. 96. ISBN 978-0-521-83222-9. Pristupljeno 10. studenoga 2011.
- ↑ Rob DeSalle; Ian Tattersall. 2008. Human origins: what bones and genomes tell us about ourselves. Texas A&M University Press. str. 146. ISBN 978-1-58544-567-7. Pristupljeno 10. studenoga 2011.
- ↑ R. J. Trent. 2005. Molecular medicine: an introductory text. Academic Press. str. 6. ISBN 978-0-12-699057-7. Pristupljeno 9. studenoga 2011.
- ↑ Donovan Webster; Spencer Wells. 20. lipnja 2010. Meeting the Family: One Man's Journey Through His Human Ancestry. National Geographic Books. str. 53. ISBN 978-1-4262-0573-6. Pristupljeno 10. studenoga 2011.
- ↑ Michael R. Speicher; Stylianos E. Antonarakis; Arno G. Motulsky. 3. veljače 2010. Vogel and Motulsky's Human Genetics: Problems and Approaches. Springer. str. 606. ISBN 978-3-540-37653-8. Pristupljeno 10. studenoga 2011.
- ↑ Dr Kutty. 14. rujna 2009. Adam's Gene and the Mitochondrial Eve. Xlibris Corporation. str. 40. ISBN 978-1-4415-0729-7. Pristupljeno 9. studenoga 2011.
- ↑ Cann RL, Stoneking M, Wilson AC. 1987. Mitochondrial DNA and human evolution. Nature. 325. (6099): 31.–36. Bibcode:1987Natur.325...31C. doi:10.1038/325031a0. PMID 3025745. Inačica izvorne stranice arhivirana 13. kolovoza 2010. Pristupljeno 3. ožujka 2014.CS1 održavanje: više imena: authors list (link)
- ↑ Gill, Victoria. 1. svibnja 2009. Africa's genetic secrets unlocked. BBC News. Pristupljeno 8. lipnja 2011. rezultati su objavljeni na internetskom izdanju časopisa Science.
- ↑ Leakey, Richard. 1994. The Origin of Humankind. Science Masters Series. Basic Books. New York, NY. str. 87.–89. ISBN 978-0-465-05313-1
- ↑ Jorde LB, Bamshad M, Rogers AR. Veljača 1998. Using mitochondrial and nuclear DNA markers to reconstruct human evolution. BioEssays. 20 (2): 126.–136. doi:10.1002/(SICI)1521-1878(199802)20:2<126::AID-BIES5>3.0.CO;2-R. PMID 9631658CS1 održavanje: više imena: authors list (link)
- ↑ Wall, J. D.; Lohmueller, K. E.; Plagnol, V. 2009. Detecting Ancient Admixture and Estimating Demographic Parameters in Multiple Human Populations. Molecular Biology and Evolution. 26 (8): 1823.–1827. doi:10.1093/molbev/msp096. PMC 2734152. PMID 19420049
- ↑ Green RE, Krause J, Briggs AW; i dr. Svibanj 2010. A draft sequence of the Neandertal genome. Science. 328 (5979): 710.–722. Bibcode:2010Sci...328..710G. doi:10.1126/science.1188021. PMID 20448178 Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: više imena: authors list (link) - ↑ ^ Reich D, Green RE, Kircher M, et al. (prosinac 2010). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature 468 (7327): 1053–60. PMID 21179161
- ↑ Reich D, Patterson N, Kircher M; i dr. Rujan 2011. Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania. Am. J. Hum. Genet. 89 (4): 516.–528. doi:10.1016/j.ajhg.2011.09.005. PMC 3188841. PMID 21944045 Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: više imena: authors list (link) - ↑ a b Laurent Abi-Rached; Jobin, M. J.; Kulkarni, S.; McWhinnie, A.; Dalva, K.; Gragert, L.; Babrzadeh, F.; Gharizadeh, B.; Luo, M.; i dr. 25. kolovoza 2011. The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans. Science. 334 (6052): 89.–94. Bibcode:2011Sci...334...89A. doi:10.1126/science.1209202. PMC 3677943. PMID 21868630. Inačica izvorne stranice arhivirana 15. travnja 2023. Pristupljeno 2. rujna 2012. Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: bot: nepoznat status originalnog URL-a (link) - ↑ a b Searching for traces of the Southern Dispersal Arhivirana inačica izvorne stranice od 10. svibnja 2012. (Wayback Machine), by Dr. Marta Mirazón Lahr, et al.
- ↑ Macaulay, V.; Hill, C; Achilli, A; Rengo, C; Clarke, D; Meehan, W; Blackburn, J; Semino, O; Scozzari, R. 2005. Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes. Science. 308 (5724): 1034.–1036. Bibcode:2005Sci...308.1034M. doi:10.1126/science.1109792. PMID 15890885
- ↑ Stanford, Craig, John S. Allen, and Susan C. Anton (Author) 2012. Biological Anthropology (2nd Edition). Prentice Hall. ISBN 0136011608 ch. 1
- ↑ a b Wood, Bernard; Richmond, Brian G. 2000. Human evolution: taxonomy and paleobiology. Journal of Anatomy. 197 (1): 19–60. doi:10.1046/j.1469-7580.2000.19710019.x. PMC 1468107. PMID 10999270CS1 održavanje: više imena: authors list (link)
- ↑ Ajit, Varki and David L. Nelson. 2007. Genomic Comparisons of Humans and Chimpanzees. Annu. Rev. Anthropol. 2007. 36:191–209: "Potvrđeno je da su razlike u sekvencama u odnosu na ljudski genom oko 1% u dijelovima koji se mogu precizno usporediti, što predstavlja razliku od oko 35 milijuna parova baza. Otkriveno je oko 45 milijuna nukleotida insercija i delecija karakterističnih za svaku vrstu, povećavajući tako razliku među genomima na oko 4%."
- ↑ Ken Sayers, Mary Ann Raghanti, and C. Owen Lovejoy. 2012. (forthcoming, october) Human Evolution and the Chimpanzee Referential Doctrine. Annual Review of Anthropology, svezak 41.
- ↑ Ruvolo, M. 1997. Genetic Diversity in Hominoid Primates. Annual Review of Anthropology, svezak 26, (1997.), str. 515.-540.
- ↑ Ruvolo, Maryellen. 1997. Molecular phylogeny of the hominoids: inferences from multiple independent DNA sequence data sets. Molecular Biology and Evolution. 14 (3): 248.–265. PMID 9066793
- ↑ Begun, David R. 2010. Miocene Hominids and the Origins of the African Apes and Humans. Annual Review of Anthropology, svezak 39: str. 67.-84.
- ↑ Wood, Bernard A. 2009. Where does the genus Homo begin, and how would we know?. Grine, Frederick E.; Fleagle, John G.; Leakey, Richard E. (eds) (ur.). The First Humans: Origin and Early Evolution of the Genus Homo. izdavač: Springer. London, UK. str. 17.–27, . ISBN 978-1-4020-9979-3CS1 održavanje: dodatna interpunkcija (link) CS1 održavanje: više imena: editors list (link) CS1 održavanje: dodatni tekst: editors list (link)
- ↑ Mitchell, Alanna. 10. siječnja 2012. DNA Turning Human Story Into a Tell-All. NYTimes. Pristupljeno 13. veljače 2012.
- ↑ Wood B. 1996. Human evolution. BioEssays. 18 (12): 945.–954. doi:10.1002/bies.950181204. PMID 8976151
- ↑ P.R. Willoughby, Paleoanthropology and the Evolutionary Place of Humans in Nature International Journal of Comparative Psychology UCLA, - 2005.
- ↑ R.D. Martin Evolution of Primates - 2002.
- ↑ [K. D. Rose -1994 DOI: 10.1002/evan.1360030505 ] - [J. Fleagle, C. Gilbert 2011-2012 [1] ] - [J. Roach 2008 [2] ] - [V. McMains - 2011 [3] ] - [2009 [4] & [5] ] [Retrieved 2012-01-01]
- ↑ Kordos L, Begun D R. 2001. Primates from Rudabánya: allocation of specimens to individuals, sex and age categories. J. Hum. Evol. 40 (1): 17.–39. doi:10.1006/jhev.2000.0437. PMID 11139358
- ↑ David W. Cameron. 2004. Hominid adaptations and extinctions. UNSW Press. str. 76. ISBN 978-0-86840-716-6. Pristupljeno 6. studenoga 2011.
- ↑ David Rains Wallace. 13. rujna 2005. Beasts of Eden: Walking Whales, Dawn Horses, and Other Enigmas of Mammal Evolution. University of California Press. str. 240. ISBN 978-0-520-24684-3. Pristupljeno 6. studenoga 2011.
- ↑ Zalmout, I. S.; Sanders, W. J.; MacLatchy, L. M.; Gunnell, G. F.; Al-Mufarreh, Y. A.; Ali, M. A.; Nasser, A.-A. H.; Al-Masari, A. M.; Al-Sobhi, S. A. 2010. New Oligocene primate from Saudi Arabia and the divergence of apes and Old World Monkeys. Nature. 466 (7304): 360.–364. Bibcode:2010Natur.466..360Z. doi:10.1038/nature09094. PMID 20631798
- ↑ Srivastava. 2009. Morphology Of The Primates And Human Evolution. PHI Learning Pvt. Ltd. str. 87. ISBN 978-81-203-3656-8. Pristupljeno 6. studenoga 2011.
- ↑ Figure 1. Phylogeny of Primate Lice from Morphological and Molecular Data Arhivirana inačica izvorne stranice od 4. prosinca 2007. (Wayback Machine), in Reed DL, Smith VS, Hammond SL, Rogers AR, Clayton DH PLoS. 2004. Genetic Analysis of Lice Supports Direct Contact between Modern and Archaic Humans. PLoS Biol. 2 (11): e340. doi:10.1371/journal.pbio.0020340. PMC 521174. PMID 15502871. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 10. ožujka 2021.CS1 održavanje: više imena: authors list (link)
- ↑ Strait DS, Grine FE, Moniz MA. 1997. A reappraisal of early hominid phylogeny. J. Hum. Evol. 32 (1): 17.–82. doi:10.1006/jhev.1996.0097. PMID 9034954CS1 održavanje: više imena: authors list (link)
- ↑ a b Bill Bryson. 2004. 28. The Mysterious Biped. A Short History of Nearly Everything. Random House, Inc. str. 522.–543. ISBN 978-0-385-66004-4
- ↑ Walker, Alan. 2006. Early Hominin Diets: Overview and Historical Perspectives. Peter Ungar (ur.). Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. Oxford University Press. US. str. 3.–10. ISBN 978-0-19-518346-7.
- ↑ Peter Ungar & Mark F. Teaford. 2002. Human Diet: Its Origin and Evolution. Bergin & Garvey. Westport, CT. str. 206. ISBN 978-0-89789-736-5
- ↑ Bogin, Barry. 1997. The evolution of human nutrition. Romanucci-Ross, Lola; Moerman, Daniel E.; & Tancredi, Laurence R. (ur.). The Anthropology of Medicine: From Culture to Method 3. izdanje izdanje. Bergen and Garvey. South Hadley MA. str. 96.–142. ISBN 978-0-89789-516-3. Inačica izvorne stranice arhivirana 3. prosinca 2003. Pristupljeno 7. ožujka 2014.CS1 održavanje: više imena: editors list (link)
- ↑ Barnicot NA. travanj/lipanj 2005. Human nutrition: evolutionary perspectives. Integr Physiol Behav Sci. 40 (2): 114.–117. doi:10.1007/BF02734246. PMID 17393680 Provjerite vrijednost datuma u parametru:
|date=(pomoć) - ↑ Leonard WR, Snodgrass JJ, Robertson ML. 2007. Effects of brain evolution on human nutrition and metabolism. Annu Rev Nutr. 27: 311.–327. doi:10.1146/annurev.nutr.27.061406.093659. PMID 17439362CS1 održavanje: više imena: authors list (link)
- ↑ "The new batch - 150,000 years ago". BBC - Science & Nature - The evolution of man.
- ↑ When humans faced extinction. BBC. 9. lipnja 2003. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 5. siječnja 2007.
- ↑ Wood, B. & Collard, M. (1999) The changing face of Genus Homo. Evol. Anth. 8(6) 195-207
- ↑ Toothy Tree-Swinger May Be Earliest Human. News. discovery.com. 21. svibnja 2010. Inačica izvorne stranice arhivirana 2. svibnja 2012. Pristupljeno 14. svibnja 2011.
- ↑ Wood B. 1999. 'Homo rudolfensis' Alexeev, 1986-fact or phantom?. J. Hum. Evol. 36 (1): 115.–118. doi:10.1006/jhev.1998.0246. PMID 9924136
- ↑ Gabounia L. de Lumley M. Vekua A. Lordkipanidze D. de Lumley H. 2002. Discovery of a new hominid at Dmanisi (Transcaucasia, Georgia). Comptes Rendus Palevol. 1 (4): 243.–253. doi:10.1016/S1631-0683(02)00032-5
- ↑ Lordkipanidze D, Vekua A, Ferring R; i dr. 2006. A fourth hominin skull from Dmanisi, Georgia. The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology. 288 (11): 1146.–1157. doi:10.1002/ar.a.20379. PMID 17031841 Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: više imena: authors list (link) - ↑ Turner W. 1895. On M. Dubois' Description of Remains recently found in Java, named by him Pithecanthropus erectus: With Remarks on so-called Transitional Forms between Apes and Man. Journal of Anatomy and Physiology. 29 (3. dio): 424.–445. PMC 1328414. PMID 17232143
- ↑ Spoor F, Wood B, Zonneveld F. 1994. Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion. Nature. 369 (6482): 645.–648. Bibcode:1994Natur.369..645S. doi:10.1038/369645a0. PMID 8208290CS1 održavanje: više imena: authors list (link)
- ↑ Bermúdez de Castro JM, Arsuaga JL, Carbonell E, Rosas A, Martínez I, Mosquera M. 1997. A hominid from the lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans. Science. 276 (5317): 1392.–1395. doi:10.1126/science.276.5317.1392. PMID 9162001CS1 održavanje: više imena: authors list (link)
- ↑ Carbonell, Eudald; Bermúdez de Castro, José M. 27. ožujka 2008. The first hominin of Europe. Nature. 452 (7186): 465.–469. Bibcode:2008Natur.452..465C. doi:10.1038/nature06815. PMID 18368116. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 26. ožujka 2008.
- ↑ Manzi G, Mallegni F, Ascenzi A. 2001. A cranium for the earliest Europeans: Phylogenetic position of the hominid from Ceprano, Italy. Proc. Natl. Acad. Sci. U.S.A. 98 (17): 10011.–10016. Bibcode:2001PNAS...9810011M. doi:10.1073/pnas.151259998. PMC 55569. PMID 11504953CS1 održavanje: više imena: authors list (link)
- ↑ Czarnetzki, A; Jakob, T; Pusch, CM. 2003. Palaeopathological and variant conditions of the Homo heidelbergensis type specimen (Mauer, Germany). Journal of Human Evolution. 44 (4): 479.–495. doi:10.1016/S0047-2484(03)00029-0. PMID 12727464
- ↑ Scientists discover hominid cranium in Ethiopia (priopćenje). Indiana University. 27. ožujka 2006. Pristupljeno 26. studenoga 2006.
- ↑ Harvati K. 2003. The Neanderthal taxonomic position: models of intra- and inter-specific craniofacial variation. J. Hum. Evol. 44 (1): 107.–132. doi:10.1016/S0047-2484(02)00208-7. PMID 12604307
- ↑ Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S. 1997. Neandertal DNA sequences and the origin of modern humans. Cell. 90 (1): 19.–30. doi:10.1016/S0092-8674(00)80310-4. PMID 9230299CS1 održavanje: više imena: authors list (link)
- ↑ Green RE; Malaspinas, Anna-Sapfo; Krause, Johannes; Briggs, Adrian W.; Johnson, Philip L.F.; Uhler, Caroline; Meyer, Matthias; Good, Jeffrey M.; Maricic, Tomislav; i dr. 2008. A complete Neandertal mitochondrial genome sequence determined by high-throughput sequencing. Cell. 134 (3): 416.–426. doi:10.1016/j.cell.2008.06.021. PMC 2602844. PMID 18692465 Eksplicitna upotreba et al. u:
|author=(pomoć) - ↑ Serre D, Langaney A, Chech M; i dr. 2004. No Evidence of Neandertal mtDNA Contribution to Early Modern Humans. PLoS Biol. 2 (3): E57. doi:10.1371/journal.pbio.0020057. PMC 368159. PMID 15024415 Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: više imena: authors list (link) - ↑ a b Viegas, Jennifer. 6. svibnja 2010. Neanderthals, humans interbred, DNA proves. Discovery News. Inačica izvorne stranice arhivirana 18. kolovoza 2010. Pristupljeno 17. kolovoza 2010.
- ↑ Gutiérrez G, Sánchez D, Marín A. 2002. A reanalysis of the ancient mitochondrial DNA sequences recovered from Neandertal bones. Mol. Biol. Evol. 19 (8): 1359.–1366. doi:10.1093/oxfordjournals.molbev.a004197. PMID 12140248CS1 održavanje: više imena: authors list (link)
- ↑ Hebsgaard MB, Wiuf C, Gilbert MT, Glenner H, Willerslev E. 2007. Evaluating Neanderthal genetics and phylogeny. J. Mol. Evol. 64 (1): 50.–60. doi:10.1007/s00239-006-0017-y. PMID 17146600CS1 održavanje: više imena: authors list (link)
- ↑ Banks, W. E; Francesco d'Errico, A. Townsend Peterson, Masa Kageyama, Adriana Sima, Maria-Fernanda Sánchez-Goñi; Peterson, A. Townsend; Kageyama, Masa; Sima, Adriana; Sánchez-Goñi, Maria-Fernanda. 2008. Harpending, Henry (ur.). Neanderthal extinction by competitive exclusion. PLoS ONE. 3 (12 e3972): e3972. Bibcode:2008PLoSO...3.3972B. doi:10.1371/journal.pone.0003972. PMC 2600607. PMID 19107186CS1 održavanje: više imena: authors list (link)
- ↑ Hortolà P, Martínez-Navarro B. 2012. The Quaternary megafaunal extinction and the fate of Neanderthals: An integrative working hypothesis. Quaternary International (in press). Bibcode:2013QuInt.295...69H. doi:10.1016/j.quaint.2012.02.037
- ↑ Mellars, Paul; Jennifer C. French. 29. srpnja 2011. Tenfold Population Increase in Western Europe at the Neandertal – to – Modern Human Transition. Science. 333 (6042): 623.–627. Bibcode:2011Sci...333..623M. doi:10.1126/science.1206930. PMID 21798948
- ↑ Brown, Terence A. 8. travnja 2010. Human evolution: Stranger from Siberia. Nature. 464 (7290): 838–839. Bibcode:2010Natur.464..838B. doi:10.1038/464838a. PMID 20376137
- ↑ a b Krause, Johannes; Fu, Qiaomei; Good, Jeffrey M.; Viola, Bence; Shunkov, Michael V.; Derevianko, Anatoli P.; Pääbo, Svante. 2010. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature. 464 (7290): 894–897. Bibcode:2010Natur.464..894K. doi:10.1038/nature08976. PMID 20336068
- ↑ Katsnelson, Alla. 24. ožujka 2010. New hominin found via mtDNA. The Scientist. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 19. ožujka 2014.
- ↑ Bokma, F.; van den Brink, V.; Stadler, T. 2012. UNEXPECTEDLY MANY EXTINCT HOMININS. Evolution. 66 (9): 2969.–2974. doi:10.1111/j.1558-5646.2012.01660.x. PMID 22946817
- ↑ Reich, David; Patterson, Nick; Kircher, Martin; Delfin, Frederick; Nandineni, Madhusudan R.; Pugach, Irina; Ko, Albert Min-Shan; Ko, Ying-Chin; Jinam, Timothy A. 2011. Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics. 89 (4): 516.–528. doi:10.1016/j.ajhg.2011.09.005. PMC 3188841. PMID 21944045
- ↑ Martinón-Torres, María; Dennell, Robin; Bermúdez de Castro, José María. 2011. The Denisova hominin need not be an out of Africa story. Journal of Human Evolution. 60 (2): 251.–255. doi:10.1016/j.jhevol.2010.10.005. ISSN 0047-2484. PMID 21129766
- ↑ Brown P, Sutikna T, Morwood MJ; i dr. 2004. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature. 431 (7012): 1055.–1061. Bibcode:2004Natur.431.1055B. doi:10.1038/nature02999. PMID 15514638 Eksplicitna upotreba et al. u:
|author=(pomoć)CS1 održavanje: više imena: authors list (link) - ↑ Argue D, Donlon D, Groves C, Wright R. 2006. Homo floresiensis: microcephalic, pygmoid, Australopithecus, or Homo?. J. Hum. Evol. 51 (4): 360.–374. doi:10.1016/j.jhevol.2006.04.013. PMID 16919706CS1 održavanje: više imena: authors list (link)
- ↑ a b Martin RD, Maclarnon AM, Phillips JL, Dobyns WB. 2006. Flores hominid: new species or microcephalic dwarf?. The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology. 288 (11): 1123.–1145. doi:10.1002/ar.a.20389. PMID 17031806CS1 održavanje: više imena: authors list (link)
- ↑ shtml Supervolcanoes, BBC2, 3. veljače 2000.
- ↑ Stanley H. Ambrose. 1998. Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans. Journal of Human Evolution. 34 (6): 623.–651. doi:10.1006/jhev.1998.0219. PMID 9650103
- ↑ Ambrose, Stanley H. 2005. Volcanic Winter, and Differentiation of Modern Humans. Bradshaw Foundation. Pristupljeno 8. travnja 2006.
- ↑ Jabr, Ferris. 18. srpnja 2012. Does Thinking Really Hard Burn More Calories?. Scientific American. Pristupljeno 23. ožujka 2013.
- ↑ Gibbons, Ann. 1998. Solving the Brain's Energy Crisis. Science. 280 (5368): 1345.–1347. doi:10.1126/science.280.5368.1345. PMID 9634409
- ↑ F. Howell; François Bourlière. 30. studenoga 2007. African Ecology and Human Evolution. Transaction Publishers. str. 398. ISBN 978-0-202-36136-9. Pristupljeno 6. studenoga 2011.
- ↑ a b c Freeman, Scott; Jon C. Herron. Evolutionary Analysis (4. izdanje.)., Pearson Education, Inc. (2007). ISBN 978-0-13-227584-2 str. 786.-788.
- ↑ Early Pleistocene third metacarpal from Kenya and the evolution of modern human-like hand morphology. doi:10.1073/pnas.1316014110 journal zahtijeva
|journal=(pomoć) - ↑ a b Plummer T. 2004. Flaked stones and old bones: Biological and cultural evolution at the dawn of technology. Am. J. Phys. Anthropol. dodatak 39: 118.–164. doi:10.1002/ajpa.20157. PMID 15605391
- ↑ Diamond, Jared. 1999. Guns, Germs, and Steel: The Fate of Human Societies. W. W. Norton. str. 39. ISBN 978-0-393-31755-8
- ↑ Ofer Bar-Yosef. The Upper Paleolithic Revolution. Annual Review of Anthropology. svezak 31, (2002.), str. 363.-393.
- ↑ April Nowell. 2010. Defining Behavioral Modernity in the Context of Neandertal and Anatomically Modern Human Populations. Annual Review of Anthropology svezak. 39: str. 437.-452. DOI: 10.1146/annurev. anthro.012809.105113
- ↑ Francesco d'Errico and Chris B. Stringer. 2011. Evolution, revolution or saltation scenario for the emergence of modern cultures? Phil. Trans. R. Soc. B 12. travnja 2011. svezak 366, broj 1567, str. 1060.-1069.
- ↑ Ambrose SH. 2001. Paleolithic technology and human evolution. Science. 291 (5509): 1748.–1753. Bibcode:2001Sci...291.1748A. doi:10.1126/science.1059487. PMID 11249821
- ↑ Mcbrearty S, Brooks AS. 2000. The revolution that wasn't: a new interpretation of the origin of modern human behavior. J. Hum. Evol. 39 (5): 453.–563. doi:10.1006/jhev.2000.0435. PMID 11102266
- ↑ Medical Research Council (UK). 21. studenoga 2009. Brain Disease 'Resistance Gene' evolves in Papua New Guinea community; could offer insights Into CJD. Science Daily (online). Science News. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 22. studenoga 2009.
- ↑ Cochran G & Harpending H. 2009. The 10,000 Year Explosion. Basic Books N.Y.
- ↑ Diamond, Jarrad (2002.), "Guns, Germs and Steel" (Penguin)
- ↑ http://www.jstor.org/discover/10.2307/2804113?uid=3738352&uid=2&uid=4&sid=21103336227741
- ↑ Are humans still evolving by Darwin's natural selection?, pristupljeno, 23. ožujka 2014.
| ||||||||||||||||||||||||||||||||||||
| ||||||||
| |||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||
